Bách khoa toàn thư cởi Wikipedia
Phiên mã nhân thực là quy trình tổ hợp RNA ở loại vật nhân thực.[1][2][3]
Bạn đang xem: quá trình phiên mã ở sinh vật nhân thực
Khái niệm này dịch kể từ thuật ngữ giờ đồng hồ Anh: eukaryotic transcription (phiên âm Quốc tế: /juːˈkærioʊt trænˈskrɪpʃən/) dùng để làm chỉ sự tổ hợp RNA dựa vào nguyên hình của DNA, ra mắt nhập tế bào của loại vật với nhân hoàn hảo hoặc loại vật nhân thực (eukaryotes).[4], [5] Đây là tên thường gọi tắt, thông thường sử dụng cho tới quy trình phiên mã xẩy ra ở group những loại vật nhân thực.
Vì những loại vật nhân thực (gồm nguyên con người) với tế bào với nhân hoàn hảo, được bảo phủ bởi vì màng nhân, tế bào lại được thường xuyên hoá cao, nên quy trình này phức tạp hơn nhiều phiên mã ở nhân sơ (xem ở trang Phiên mã). Thêm nhập cơ, phiên mã ra mắt bên trên DNA khuôn, nhưng mà DNA đó lại trực thuộc bào quan tiền đặc biệt quan trọng gọi là NST với cấu tạo phức tạp, nên quy trình chỉ ra mắt Khi trước cơ (vào tiến độ chuẩn chỉnh bị) với sự toá xoắn NST, toá xoắn DNA và dãn xoắn-tách mạch gen nguyên hình, tức thị tương quan cả cho tới điều hoà gen. Dường như, sau khoản thời gian RNA và được tổ hợp đi ra thì vẫn phải qua chuyện quy trình chế biến đổi mới nhất trở nên thành phầm với tính năng sinh học tập hoàn hảo.
Tổng quan[sửa | sửa mã nguồn]
- Phiên mã (transcription) về thực chất là quy trình tổ hợp RNA kể từ mạch khuôn của gen. Trong quy trình này, chuỗi pôlyđêôxyribônuclêic của gen bên trên DNA được sản xuất khuôn, nhằm tổ hợp nên chuỗi mới nhất, tuy nhiên lại là chuỗi pôlyribônuclêic của RNA. Vì gen là đoạn xác lập của DNA với 2 mạch, tuy nhiên chỉ 1 mạch cố định và thắt chặt được lựa chọn thực hiện khuôn sẽ tạo đi ra phân tử RNA, còn mạch cơ là mạch ko cần khuôn, nên mạch thực hiện khuôn được gọi là mạch mã gốc.[6] Trong Di truyền học tập phân tử, mạch mã gốc này là mạch đối nghĩa.[7], [8]. Phân tử RNA được tổ hợp với trình tự động những ribônuclêôtit bổ sung cập nhật cho tới trình tự động đêôxyribônuclêôtit ở mạch đối nghĩa của gen theo gót phép tắc bổ sung cập nhật, vì thế thẳng đem những cỗ thân phụ mã DT, hoặc sách VN thông thường gọi là những côđon.
- Quá trình này ra mắt nhập nhân của tế bào và tổ chức qua chuyện thân phụ tiến độ tuần tự động tương tự động như phiên mã nhân sơ: chính thức, kéo dãn dài và kết cổ động (xem ở trang phiên mã). Tuy nhiên, khối hệ thống những nhân tố nhập cuộc phiên mã thật nhiều và phức tạp hơn nhiều ở nhân sơ.[9] Các điểm đa số như sau.
- Vì phiên mã ra mắt nhập nhân, nhưng mà nhân với màng bảo phủ, nên phiên mã ko thể ra mắt mặt khác cùng theo với dịch mã như ở nhân sơ.
- Ở vi trùng (nhân sơ), phiên mã của toàn bộ những loại gen tạo nên nhiều loại RNA không giống nhau hầu hết chỉ được xúc tác bởi vì một loại RNA pôlymêraza;[3] còn ở nhân thực, với tối thiểu thân phụ loại RNA pôlymêraza không giống nhau, gọi tắt là Pol I, Pol II và Pol III nhập tổ hợp những loại đa số là mRNA, tRNA và rRNA, chưa tính nhiều loại RNA không giống nữa (xem ở trang Danh sách RNA).
• Pol I (RNA pôlymêraza 1) xúc tác phiên mã của toàn bộ những gen mã hoá RNA ribôxôm (rRNA) nước ngoài trừ 5S. Những gen rRNA này được tổ chức triển khai trở nên một đơn vị chức năng phiên mã độc nhất và được phiên trở nên một mặt hàng liên tiếp. Phân tử nguyên sơ sau này được xử lý trở nên thân phụ loại: 18S, 5,8S và 28S. Sự phiên mã gen rRNA ra mắt nhập hoạnh họe nhân (nucleolus hoặc nhân con) kể từ cơ kết phù hợp với những prôtêin nhằm tạo hình những ribôxôm.[10], [11]
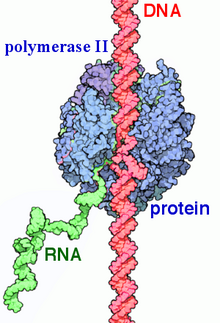
• Pol II (RNA pôlymêraza 2 - hình 2) với tầm quan trọng chủ yếu nhập xúc tác phiên mã hàng nghìn gen mã hóa prôtêin trở nên RNA vấn đề.[9]
• Pol III (RNA pôlymêraza 3) xúc tác phiên mã một trong những lượng chắc chắn những gen mã hóa RNA vận gửi (tRNA). Pol III còn xúc tác tổ hợp RNA ko mã hóa độ dài rộng nhỏ khác ví như rRNA 5S, SNRNA, SRP RNA, RNA ribonuclease.[12]
- Quá trình phiên mã ra mắt nhập nhân tế bào, tạo nên những RNA nguyên sơ hoặc cũng gọi là chi phí RNA (pre RNA). Sau cơ, những RNA nguyên sơ này cần trải qua chuyện tiến độ chế biến đổi (processing) ở nhập nhân, tạo ra RNA trưởng thành và cứng cáp, rồi vừa được "xuất khẩu" kể từ nhân thanh lịch tế bào hóa học nhằm triển khai tính năng sinh học tập của bọn chúng (hình 3).
- Trong quy trình chế biến đổi mRNA nguyên sơ sẽ tạo đi ra RNA trưởng thành và cứng cáp, vì như thế gen nhân thực nằm trong loại gen phân miếng, bao gồm những đoạn intrôn (không với mã) đan xen với những đoạn êxôn (có mã), nên quy trình chế biến đổi mRNA nguyên sơ còn cần phải cắt vứt những đoạn ko mã (intrôn) rồi phân giải bọn chúng, mặt khác ghép nối những đoạn với mã (êxôn) lại trở nên cấu tạo tuyến tính hoàn hảo chứa chấp những trình tự động ribônuclêôtit liên tiếp nhằm dịch mã nhanh gọn và đúng đắn.

- Tóm tắt những loại Pol như bảng sau:
| Tên | Sản phẩm |
| RNA pôlymêraza 1 (Pol I, Pol A) | RNA ribôxôm (rRNA) rộng lớn bao gồm rRNA 28S, 18S và 5,8S. |
| RNA pôlymêraza 2 (Pol II, Pol B) | RNA vấn đề (mRNA), RNA nhân nhỏ (snRNA), RNA siêu nhỏ (microRNA) |
| RNA pôlymêraza 3 (Pol III, Pol C) | RNA vận gửi (tRNA) và một trong những loại khác ví như rRNA 5s, srpRNA, RNA nhận dạng tín hiệu (SRP RNA). |
Cơ chế phiên mã[sửa | sửa mã nguồn]
Yếu tố phiên mã[sửa | sửa mã nguồn]
Như bên trên vẫn nói: Tại nhân sơ, phiên mã tự duy nhất loại enzym đảm nhận gọi là RNA pôlymêraza. Enzym này còn có kĩ năng phát động phiên mã ngay lập tức, sau khoản thời gian nó gắn bên trên một điểm của gen gọi là vùng phát động (promoter). Nhưng ở nhân thực, cần nhận thêm một bước nữa tổ chức trước, nhập cơ enzym này chỉ hoàn toàn có thể gắn kèm với promoter với việc trợ hùn của tinh vi prôtêin đặc thù gọi là những nhân tố phiên mã hoặc tinh vi trợ hùn phiên mã (hình 2). Phức thích hợp này là 1 trong những phần luôn luôn phải có của máy bộ phiên mã cho tới ngẫu nhiên gen nào là nhập tế bào nhân thực. Cách sẵn sàng này là tiến độ sẵn sàng cho tới khởi điểm phiên mã, nên gọi là chi phí khởi điểm (pre-initiation).[13] Chi tiết về tinh vi này coi ở trang Phức thích hợp chi phí khởi điểm phiên mã (Transcription preinitiation complex).
- Vùng phát động phiên mã của gen nhân thực (promoter eukaryotic) to hơn và phức tạp rất là nhiều đối với của vi trùng (promoter prokaryotic) tuy nhiên cả nhì đều sở hữu một trình tự động TATA... gọi tắt là hộp TATA (TATA box). Ví dụ, nhập gen tổ hợp thymidine kinase của loài chuột, vỏ hộp này và đúng là TATAAAA, xác định bên trên toạ chừng -30 đối với địa điểm chính thức (+1), được phát âm theo phía 5'-3' bên trên mạch bổ sung cập nhật (nontemplate).[14]
- Pol II - tự động nó - trọn vẹn đầy đủ kĩ năng nhằm xúc tác tổ hợp RNA dựa vào khuôn DNA, tuy nhiên nó ko thể tự động nhận thấy thẳng vùng phát động (promoter) này của gen nhưng mà nó sẽ bị xúc tác. Do cơ, nó cần thiết nhiều nhân tố trợ hùn, tạo nên trở nên một hội tụ những phân tử phức tạp và đặc thù tương quan cho tới nhau, gọi là phức thích hợp phát động phiên mã Pol II (Pol II transcription preinitiation complex - ghi chép tắt: PIC-Pol II). Trong từng PIC (viết tắt kể từ preinitiation complex) của Pol II, có rất nhiều phân tử không giống nhau gọi là nhân tố hoặc thành phần (element). Mỗi nhân tố vào vai trò như là 1 trong những địa điểm kết nối cho những bộ phận ví dụ không giống nhập "bộ máy" phiên mã công cộng, chứ không hề cần toàn bộ từng loại phiên mã, nên gọi là những nhân tố phiên mã công cộng (general transcription factors). Các nhân tố phiên mã công cộng này bao hàm TFIIB, TFIID, TFIIE, TFIIF và TFIIH (hình 4). Sau trên đây, gọi tắt Pol II transcription preinitiation complex là PIC cho tới giản dị.
Tiền khởi điểm phiên mã[sửa | sửa mã nguồn]
- Đầu tiên, PIC được tạo nên nhờ prôtêin link TATA với TBP, một tè đơn vị chức năng của TFIID. Liên kết này với TATA thực hiện cho tới DNA bị uốn nắn cong lại ở điểm xác lập (hình 5). Tiếp theo gót, TFIIA rồi cho tới TFIIB link với tinh vi DNA-TBP ở cả thượng lưu (phía trên) và hạ lưu (phía dưới) của vỏ hộp TATA. Phức thích hợp DNA-TBP-TFIIB tồn tại vẫn hoàn toàn có thể link với Pol II, sẽ tiến hành TFIIF gửi cho tới vùng phát động. Rồi TFIIE link hâu phương Pol II và như 1 ĐK tiên quyết cần thiết liên kết với TFIIH. Lúc này, PIC được xem là và được lắp đặt ráp hoàn hảo.[15]
- Trong tình huống không tồn tại vỏ hộp TATA, thì tinh vi PIC vẫn hoàn toàn có thể chính thức hoạt động và sinh hoạt bằng phương pháp gắn những tè đơn vị chức năng TAF của TFIID với những nhân tố không giống. Trường thích hợp này sẽ không tế bào miêu tả ở trên đây. Tập thích hợp PIC được kích hoạt nhập phản xạ với hóa học kích hoạt gắn kèm với nhân tố đẩy mạnh (enhancer) tương tự chuỗi hoạt hóa ở nấm men, điều này tiếp tục thay cho thế những tinh vi đồng kích hoạt (coactivator) bao hàm những hóa học thay đổi nhiễm sắc tử (chromatine) và nhân tố trung gian dối (mediator) tương tác thẳng với Pol II và những nhân tố phiên mã công cộng.[16]
- Ở tình huống với vỏ hộp TATA, cuối tiến độ phát động này, thì PIC và địa điểm của chính nó được tế bào miêu tả ở hình 6.
- Tóm lại, sự lắp đặt ráp những nhân tố phiên mã ra mắt theo gót quá trình công cộng như sau:
- TBP link với vỏ hộp TATA (TBP là 1 trong những tè đơn vị chức năng của TFIID).
- TBP tương tác với TFIIA, nhờ cơ TFIIA lắp đặt nhập vùng phát động.
- Xuất hiện nay tương tác TBP với TFIIB, nhờ cơ TFIIB lắp đặt nhập vùng phát động.
- TFIIB tương tác với Pol II và tương tác TFIIB với TFIIF hỗ trợ cho Pol II và TFIIF nhập vùng phát động.
- TFIIE nhập cuộc nhập tinh vi và "kết nạp" TFIIH với hoạt tính kinase, nó phôtphoryl hoá Pol II ở CTD, và hoạt động và sinh hoạt hêlicaza dãn xoắn và tách mạch gen bên trên vùng phát động. Đồng thời, nó cũng "kết nạp" những prôtêin sửa chữa thay thế nhập phiên mã.
- Các tè đơn vị chức năng của TFIIH phát động tính năng ATPase và hêlicaza thay đổi sắc thái của gen được phiên mã trở nên dạng tuyến tính.
- Tháo xoắn trọn vẹn đoạn gen chuẩn bị phiên mã, tạo nên trở nên một cấu tạo gọi là "bóng phiên mã" (hình 1).
- Bóng phiên mã tương tác với Pol II, kể từ cơ quy trình tổ hợp RNA phi vào tiến độ 1 (khởi đầu).
-

Xem thêm: đạo hàm của căn x
Hình 4: Sơ thiết bị hội tụ PIC bao gồm những nhân tố phiên mã công cộng và Pol II.
-

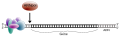
Hình 5: Mô hình giản dị khởi điểm phiên mã nhân thực. 1 = Vị trí chính thức phiên mã. 2 = Hộp TATA với Pol II vẫn gắn kèm với PIC. 3 = Trình tự động đẩy mạnh link prôtêin kích hoạt.
-
Hình 6: Sơ thiết bị sự kết nối của Pol II với tinh vi PIC cuối tiến độ "khởi động" phiên mã.
Diễn biến đổi phiên mã[sửa | sửa mã nguồn]
1) Giai đoạn khởi điểm (initiation)[sửa | sửa mã nguồn]
Khi "bóng phiên mã" tương tác với Pol II (bước 8 ở trên) thì cũng chính là tiến độ khởi điểm phiên mã được tổ chức. Lúc này tạo hình nhập "bụng" của Pol II một cấu tạo "phân tử lai" trong thời điểm tạm thời xuất hiện: DNA-RNA, kể từ cơ những ribônuclêôtit được lắp đặt nhập khuôn theo gót phép tắc bổ sung cập nhật (A-U, G-X). Rồi tiến độ kéo dãn dài ra mắt ngay lập tức.[3][17][18]
Xem thêm: cách tính hiệu suất phản ứng
2) Giai đoạn kéo dãn dài (elongation)[sửa | sửa mã nguồn]
- Sau Khi tổ hợp được một quãng cộc ribônuclêôtit (khoảng rộng lớn 10 base), thì Pol II bay ngoài vùng phát động tương đương bay ngoài PIC nhằm tự động phiên mã nốt đoạn còn sót lại của gen. Trong tiến độ này, kể từ tổng hợp "phân tử lai" rằng bên trên xuất hiện nay 2 sợi rời khỏi theo gót 2 kênh riêng biệt biệt: sợi RNA càng ngày càng nhiều năm (hình 2), còn sợi DNA (khuôn) tiếp tục tái ngắt kết phù hợp với sợi DNA bổ sung cập nhật với nó ở hâu phương Pol II, mặt khác đóng góp xoắn lại ngay lập tức sau khoản thời gian và được Pol II "đọc" hoàn thành. Giai đoạn này rằng công cộng tương tự động như ở loại vật nhân sơ (vi khuẩn).
- Mặc cho dù trình diễn biến đổi công cộng của tiến độ kéo dãn dài ở nhân thực tương tự ở vi trùng, tuy nhiên có rất nhiều phân tách khác lạ. Một trong những khác lạ chủ yếu là cần phải có những nhân tố kéo dãn dài (elongation factors) với tầm quan trọng kích ứng sự kéo dãn dài phiên mã, ví như P-TEFb đặc biệt quan trọng cần thiết. P-TEFb phôtphoryl hoá Ser-2 và kích hoạt SPT5 tương đương TAT-SF1. Trong số đó, SPT5 là 1 trong những nhân tố phiên mã hùn phối hợp enzym 5'-capping nhập Pol II.[19]
3) Giai đoạn kết cổ động (termination)[sửa | sửa mã nguồn]
- Khi Pol II trượt cho tới một cấu tạo gọi là yếu tố kết thúc (transcription terminator) ở cuối gen cần thiết phiên, thì nó sẽ bị tách ngoài đoạn DNA nguyên hình này và quy trình phiên mã cho một gen của chính nó kết cổ động. Mỗi phiên phiên mã, Pol II chỉ phiên được một mRNA chứa chấp 1 gen, khác hoàn toàn với ở nhân sơ là mRNA nhân sơ bao gồm 1 bạn dạng phiên tuy nhiên lại chứa chấp vấn đề của rất nhiều gen (cụm gen ở operon). Đừng sai sót nhân tố kết cổ động này với cỗ thân phụ kết cổ động (Stop codon) ở gen và cũng chính là của mRNA với cấu tạo và tầm quan trọng khác hoàn toàn nhập dịch mã.
- Xem cụ thể rộng lớn về yếu tố này ở trang Yếu tố kết cổ động phiên mã.
Bởi vì như thế gen cấu tạo của nhân thực bao gồm nhiều đoạn êxôn (có mã) lẫn lộn đối với cả intrôn (không mã), nên quy trình phiên mã nhân thực kết cổ động nhưng mà mới nhất chỉ tạo nên RNA nguyên sơ (hay chi phí RNA - primordial RNA) chưa xuất hiện tính năng sinh học tập. Do cơ, thành phầm nguyên sơ này còn cần qua chuyện quy trình chế biến đổi hoặc xử lý (RNA processing) mới nhất tạo ra RNA trưởng thành và cứng cáp.
Chế biến đổi RNA[sửa | sửa mã nguồn]
Quá trình chế biến đổi RNA được phân phát hiện nay khoảng chừng từ thời điểm năm 1977-1978. Quá trình này còn được gọi là xử lý RNA, bao gồm 3 thay đổi chính:
- Gắn chóp (mũ) 7-mêtyl-guanylat nhập đầu 5' của RNA.
- Thêm đuôi pôlyA nhập đầu 3' của RNA.
- Cắt-nối (splicing) bao gồm hạn chế vứt intrôn (không mã), tích lại êxôn (có mã) rồi tiếp nối những êxôn vẫn hạn chế cùng nhau theo như đúng trình tự động nó vốn liếng với bên trên gen nguyên hình.[2], [3]
Xem cụ thể rộng lớn ở trang Xử lý RNA.
Xem thêm[sửa | sửa mã nguồn]
- Phiên mã
- Xử lý RNA
Nguồn trích dẫn[sửa | sửa mã nguồn]
- ^ Krishnamurthy & Michael Hampsey. “Eukaryotic transcription initiation Shankarling”.
- ^ a b Campbell và nằm trong sự: "Sinh học" - Nhà xuất bạn dạng giáo dục và đào tạo, 2010.
- ^ a b c d Phạm Thành Hổ: "Di truyền học" - Nhà xuất bạn dạng giáo dục và đào tạo, 1998.
- ^ “Stages of transcription”. Khan Academy. Truy cập 7 mon 12 năm 2018.
- ^ “Transcription in Eukaryotes Genetics”. Biology Discussion. Truy cập 7 mon 12 năm 2018.
- ^ "Sinh học tập 12" - Nhà xuất bạn dạng giáo dục và đào tạo, 2019
- ^ Vicent Pelechano & Lars M. Steinmetz. “Gene regulation by antisense transcription”.
- ^ “Medical Definition of Antisense”. Bản gốc tàng trữ ngày 9 mon 12 năm 2018.
- ^ a b Shankarling Krishnamurthy & Michael Hampsey. “Eukaryotic transcription initiation”.
- ^ Sirri, Valentina; Silvio Urcuqui-Inchima; Pascal Roussel; Danièle Hernandez-Verdun (2008). “Nucleolus: the fascinating nuclear body”. Histochem Cell Biol. 129 (1): 13–31. doi:10.1007/s00418-007-0359-6. PMC 2137947. PMID 18046571.
- ^ Fromont-Racine, Micheline; Senger, Bruno; Saveanu, Cosmin; Fasiolo, Franco (tháng 8 năm 2003). “Ribosome assembly in eukaryotes”. Gene. 313: 17–42. doi:10.1016/S0378-1119(03)00629-2.
- ^ Dieci, Giorgio; Fiorino, Gloria; Castelnuovo, Manuele; Teichmann, Martin; Pagano, Aldo (tháng 12 năm 2007). “The expanding RNA polymerase III transcriptome”. Trends in Genetics. 23 (12): 614–622. doi:10.1016/j.tig.2007.09.001. PMID 17977614.
- ^ “Transcription factors”. Khan Academy. Truy cập 7 mon 12 năm 2018.
- ^ “15.3: Eukaryotic Transcription”.
- ^ Donal S Luse. “The RNA polymerase II preinitiation complex”.
- ^ “Current Biology”.
- ^ Benjamin L. Allen and Dylan J. Taatjes. “The Mediator complex: a central integrator of transcription”.
- ^ Đỗ Lê Thăng: "Di truyền học" - Nhà xuất bạn dạng giáo dục và đào tạo, 2003
- ^ Abbie Saunders, Leighton J. Vi xử lý Core & John T. Lis. “Breaking barriers to lớn transcription elongation”.











Bình luận